1. 서 론
영남지역을 관통하는 낙동강은 이 지역의 최대 수자원 중 하나로 유로연장은 525 km이고 유역면적은 23,859 km2에 달한다[1-3]. 낙동강은 부산광역시, 대구광역시, 울산광역시, 경상북도, 경상남도 지역 약 1,000만 인구의 식수원임에도 불구하고 유역 내 도시의 하・폐수처리시설 방류수 및 비점오염원이 유입되고 있어 수질관리가 중요하다. 1987년 완공된 낙동강 하구언으로 인하여 물 흐름이 정체되는 현상이 발생하여 수위 및 수질 관리에 어려움이 있었으며, 2008~2013년에 걸쳐 건설된 8개의 보(상주보, 낙단보, 구미보, 칠곡보, 강정고령보, 달성보, 합천창녕보, 창녕함안보)로 인해 물의 체류시간이 증가하여 낙동강의 환경에 녹조현상 심화 등 여러 가지 변화가 나타났다[4,5].
녹조현상은 일차 생산자인 조류(algae)가 서식하기 좋은 환경이 갖춰지며 대발생하여 하천과 호소를 뒤덮는 현상을 말한다. 특히 여름~초가을 높은 온도에서 주로 서식하는 남조류가 진한 녹색을 가지기 때문에 녹조현상으로 불린다. 남조류의 대표종인 Microcystis는 남극을 제외한 모든 대륙에서 발생하고 약 108개국에서 Microcystis 발생에 관한 문헌이 조회될 정도로 녹조는 전 세계적으로 발생하는 문제이다[6]. 녹조의 발생은 심미적 문제, 생태계 파괴, 정수처리 공정에 악영향 등 다양한 피해를 준다. 남조류 중 일부 종은 Geosmin과 2-MIB로 대표되는 냄새유발물질을 내놓고 일부 종은 마이크로시스틴(microcystin) 등 독성물질도 내놓는다. 정수처리시설에서 냄새유발물질 및 독성물질을 제거하기 위해서는 오존 공정, 활성탄 공정 등 고도처리공정이 필요하며 환경 비용이 증가한다.
환경부는 녹조현상을 예방하고 국민에게 올바른 정보를 제공하고자 1998년부터 조류경보제를 운영하였고 2012년에는 조류경보제의 지점이 아닌 4대강 유역에서의 조류를 감시하고자 수질예보제를 시행하였다. 조류경보제는 조류 발생상황을 주기적으로 모니터링하고 필요시 경보를 발령하여 수돗물을 안정적으로 공급하기 위해 하천과 호소 28개 지점에서 수온, pH, 용존산소(DO), 클로로필-a, 남조류 세포수 등의 항목을 주 1회 이상 측정하여 공개하는 제도이다. 조류 세포수가 관심(1,000~10,000 cells/mL), 경계(10,000~1,000,000 cells/mL), 조류 대발생(1,000,000 cells/mL 이상), 해제(1,000 cells/mL 미만)의 기준으로 2회 연속 기준을 초과하였을 시 해당 단계를 발령한다. 수질예보제는 4대강의 16개의 보 중 조류경보제 시행 지점인 4개 보를 제외한 12개의 보에서 공공수역의 사전예방적 수질 관리를 위하여 시행했던 제도로 기상, 수질, 유량을 분석하여 7일간의 수질변화를 예측한 결과를 주 2회(월, 목) 예보하였다[7]. 그러나 조류경보제와 수질예보제가 남조류 세포수를 기반으로 경보를 발령하며 발령 기준이 비슷하여 헷갈린다는 지적이 이어져왔다. 이에 환경부는 다년간에 걸친 분석 끝에 2020년 5월부터 조류경보제와 수질예보제를 통합하여 운영 중이다. 조류경보제는 신규 지점인 낙동강 물금・매리 지점을 추가하였고 기존 수질예보제 지점은 모두 조류경보제의 조류관찰지점으로 표기하였다. 조류경보제의 경우 직관적인 발령 기준(1 mL당 유해남조류 세포수 1천, 1만, 10만, 100만)으로 통일하여 각 단계의 기준을 1회 초과시 발령, 2회 미만으로 유지시 해제하는 것으로 단순화하였고 조류 관찰지점의 경우 경보를 발령하지 않는다.
이처럼 환경부는 매년 발생하는 녹조현상에 유연하게 대처하기 위해 조류경보제와 수질예보제를 운영해왔고 2020년에 들어서는 수질예보제를 조류경보제와 통합하여 운영하는 등 조류로부터의 피해를 예방하고 국민에게 관련 정보제공과 안전한 수자원 확보를 위해 노력해 왔다. 본 연구에서는 이러한 노력에도 불구하고 매년 녹조가 대량 발생하여 문제를 겪고 있는 낙동강 유역 중 대구경북에 속하는 지점의 녹조 발생현황을 수질예보제 시행 직후인 2012년부터 수질예보제의 조류경보제 통합 직전인 2019년까지 기간으로 알아보고 조류경보제와 수질예보제에 관해 상호 비교하고자 한다.
2. 연구방법
2.1. 대상지점 선정
본 연구는 조류경보제와 수질예보제 시행 지점 중 대구・경북 지역에 포함되는 17개 지점을 분석 대상으로 하였다. 수질예보제에 해당하는 낙동강의 보 지점은 상주보(ND-1), 낙단보(ND-2), 구미보(ND-3), 칠곡보(ND-5), 강정고령보(ND-7), 달성보(ND-8)의 6개소로, 각 보를 기준으로 상류 500 m가 측정지점이다. 조류경보제 지점은 상수원으로 활용되는 구간으로 공산지 중앙부(WS-1), 공산지 취수탑(WS-2), 칠곡보 상류 22 km (ND-4), 강정고령보 상류 7 km (ND-6), 덕동호 댐(WS-3), 안계호 취수탑(WS-4), 영천호 취수탑(WS-5), 운문호 댐(WS-6), 운문호 취수탑(WS-7), 진전지 상류(WS-8), 진전지 하류(WS-9) 등 11개소이다.
수집된 자료의 결과 표기는 수질예보제와 조류경보제로 구분하지 않고 낙동강 본류에 포함된 6개의 보 지점과 낙동강의 상수원 2개 지점(강정고령보 상류 7 km, 칠곡보 상류 22 km)을 묶어 낙동강 지점(ND-1~ND-8)으로 표현하고 나머지 조류경보제 9개 지점을 묶어 상수원 지점(WS-1~WS-9)으로 표현하였다.
2.2. 자료 수집 및 분석
자료 분석을 위해 국립환경과학원이 운영 중인 ‘물환경정보시스템(water.nier.go.kr)’에서 낙동강 수계 중 대구・경북 지역에 포함되는 지점(보 6개, 상수원 11개 지점)에 대해 자료를 수집하여 활용하였다. 수집항목은 지점별 조사일, 남조류 세포수, 우점종, 클로로필-a, 수온 등이다. 우점종 자료의 경우 조류경보제는 유해남조류 4종(Anabeana, Aphanizomenon, Microcystis, Oscillatoria)별 세포수가 개별적으로 제시되어 있으나, 수질예보제는 유해남조류의 우점종만 표기되어 있다. 측정주기는 기본적으로 주 1회 이상이나 조류 발생 정도에 따라 주 2~3회의 측정이 이루어졌다. 수집된 자료의 기간은 2012~2019년 사이이었으나, 조류경보제 지점 중 칠곡보 상류 22 km (ND-4), 강정고령보 상류 7 km (ND-6) 지점은 2016년부터, 공산지 중앙부(WS-1), 공산지 취수탑(WS-2) 지점은 2014년부터, 진전지 상류(WS-8), 진전지 하류(WS-9) 지점은 2013년부터 각각 측정이 시작되어 해당 지점들은 시작하는 년도부터의 자료를 수집하였다.
조류 및 수질 항목간 상관분석은 SPSS로 하였으며 피어슨 상관계수(Pearson correlation coefficient)를 구하였다. 피어슨 상관계수는 X, Y 두 인자의 상관성을 –1에서 1 사이의 값으로 나타내주는 계수로서 r값이 1에 근접할수록 양의 상관관계를 보이고 –1에 근접할수록 음의 상관관계를 보이며 0에 가까우면 상관성이 없다고 판단한다. 남조류의 발생에는 다양한 인자가 복잡적인 작용을 통해 만들어진 환경의 영향이 작용한다. 다만 이번 연구의 특성상 기존에 조류경보제와 수질예보제를 통해 공개되는 자료만을 활용한 결과이기에 여러 연구를 통해 밝혀진 유해남조류 발생과 연관성이 있는 인자의 복합적인 상관성을 유추하기에는 무리가 있다고 판단하여 공개되는 인자인 pH, 수온, DO, Chl-a와 유해남조류 발생량 간 각각의 연관성만을 확인하기 위해 Pearson 상관계수를 사용하였다.
3. 결과 및 고찰
3.1. 유해남조류 발생 현황
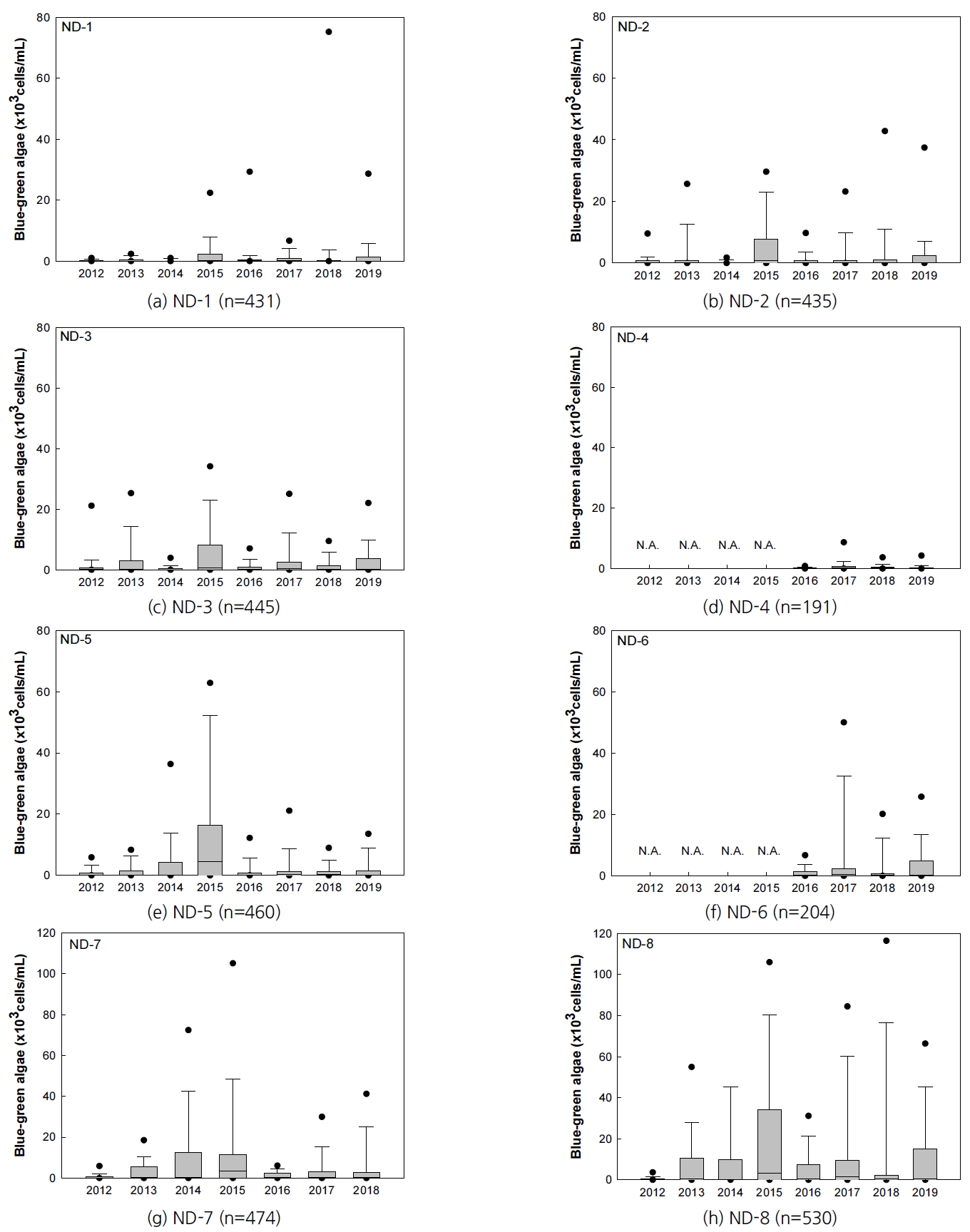
2012년부터 2019년 사이 대구・경북지역 낙동강 지점의 월 평균 유해남조류 발생량을 Fig. 1에 정리하였다. 낙동강 지점의 월별 유해남조류 발생량은 보가 완공되고 난 직후인 2012년 월평균 10,000 cells/mL을 넘지 않았으나 2013년 최대 30,326 cells/mL, 2014년 최대 83,762 cells/mL, 2015년 최대 98,888 cells/mL로 점차 증가했다가 2016년부터 감소하는 추세이다. 매년 여름철(6~8월)의 유해남조류 발생량이 높았으며 낙동강의 하류 지점으로 갈수록 유해남조류 발생 정도가 심해지는 것을 볼 수 있다.
대구・경북 지역 낙동강 지점(ND-1~ND-8) 및 상수원 지점(WS-1~WS-9)의 2012~2019년 사이 녹조 발생현황을 알아보기 위해 수집한 데이터를 이용하여 각 지점별로 연도별 유해남조류 발생량을 분석하였다(Fig. 2, 3). 낙동강 지점(ND-1~ND-8)은 Fig. 2의 (a)~(h)에 보의 위치에 따라 낙동강 상류부터 하류 순서로 나타내었다. 2012년부터 2019년 사이 대구・경북 지역의 낙동강 지점에서 측정된 유해남조류의 최대치는 495,360 cells/mL이었다. 각 지점별 유해남조류 발생량의 최대값은 상주보(ND-1) 90,263 cells/mL, 낙단보(ND-2) 89,200 cells/mL, 구미보(ND-3) 79,499 cells/mL, 칠곡보 상류 22 km (ND-4) 24,742 cells/mL, 칠곡보(ND-5) 300,500 cells/mL, 강정고령보 상류 7 km (ND-6) 51,561 cells/mL, 강정고령보(ND-7) 261,219 cells/mL, 달성보(ND-8) 495,360 cells/mL로 낙동강 하류로 갈수록 증가하는 경향을 보였다. 다만, ND-4와 ND-6지점은 2012~2015년 사이 자료가 없어 자료 분석 및 단순 비교의 한계가 있다.
각 지점별 유해남조류 발생량의 평균은 상주보(ND-1) 1,771 cells/mL, 낙단보(ND-2) 3,159 cells/mL, 구미보(ND-3) 2,931 cells/mL, 칠곡보 상류 22 km (ND-4) 632 cells/mL, 칠곡보(ND-5) 4,857 cells/mL, 강정고령보 상류 7 km (ND-6) 3,565 cells/mL, 강정고령보(ND-7) 6,875 cells/mL, 달성보(ND-8) 12,678 cells/mL로 하류부인 ND-5~ND-7지점의 발생량 평균이 상류부인 ND-1~ND-4지점의 발생량 평균보다는 높게 나타났다. 다만, 앞서 언급한 것과 같이 ND-4와 ND-6지점은 2012~2015년 사이 자료가 없어 자료 분석 및 단순 비교의 한계가 있다.
2016~2019년 사이 자료를 분석하면 칠곡보(ND-5) 지점의 유해남조류 평균 발생량은 칠곡보 상류 22 km (ND-4) 지점에 비해 약 2.9배 높고, 강정고령보(ND-7) 지점의 유해남조류 평균 발생량은 강정고령보 상류 7 km (ND-6) 지점에 비해 약 1.5배 높아 보에 근접할수록 유해남조류 발생량이 증가했다. 이는 보에 인접한 지점에서 유속 감소에 의한 영향으로 조류발생이 증가했기 때문으로 판단된다[8].
낙동강 지점의 유해남조류 발생은 계절적으로 볼 때 5~10월에 집중되고 12~2월 겨울철에는 대부분 지점에서 발생하지 않았다. 이는 유해남조류의 발생이 상하류 및 보에서의 거리에 관계없이 계절적 영향을 크게 받는다는 것을 나타낸다. 수온이 유해남조류 발생에 미치는 영향은 3.3절에서 보다 자세하게 설명되어 있다.
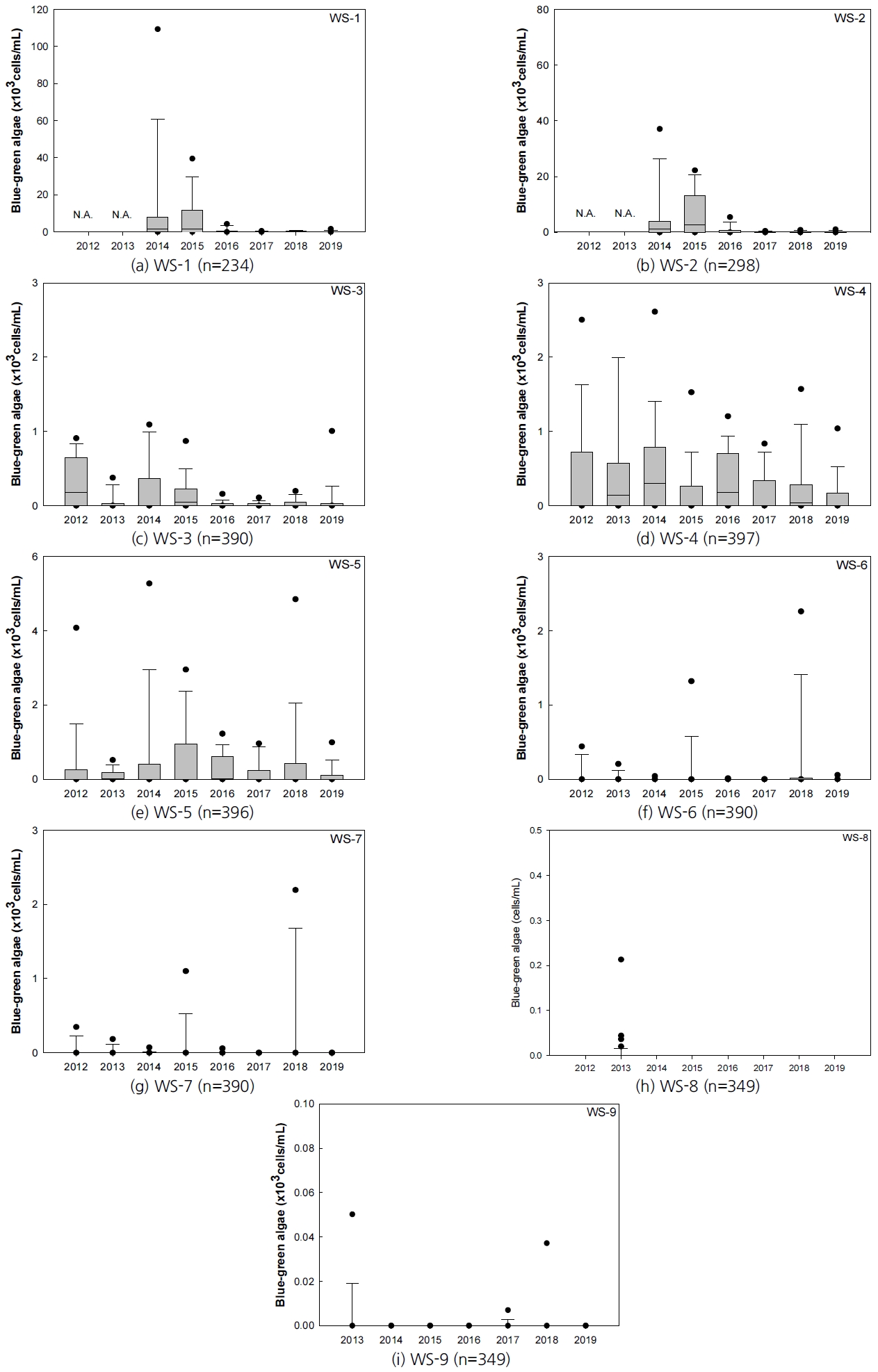
상수원 지점(WS-1~WS-9)의 유해남조류 발생 현황은 Fig. 3의 (a)~(i)에 정리하였다. 상수원 지점의 유해남조류 발생은 낙동강 본류 지점과 비교해서 상당히 적은 수치를 보인다. 이는 상수원 지점(WS-1~WS-9)인 공산지, 덕동호, 안계호, 영천호, 운문호, 진전지는 모두 상수원 보호구역으로 지정되어 있어 오수, 분뇨, 가축분뇨 등 수질 오염물질의 유입이 적었기 때문으로 판단된다. 2012년부터 2019년 사이 대구・경북 지역의 상수원 지점에서 측정된 유해남조류의 최대치는 112,000 cells/mL이었다.
각 지점별 유해남조류 발생량의 최대값은 공산지 중앙부(WS-1) 112,000 cells/mL, 공산지 취수탑(WS-2) 40,250 cells/mL, 덕동호 댐(WS-3) 1,582 cells/mL, 안계호 취수탑(WS-4) 6,733 cells/mL, 영천호 취수탑(WS-5) 9,748 cells/mL, 운문호 댐(WS-6) 3,320 cells/mL, 운문호 취수탑(WS-7) 3,801 cells/mL, 진전지 상류(WS-8) 213 cells/mL, 진전지 하류(WS-9) 222 cells/mL로 나타났다. 각 지점별 유해남조류 발생량의 평균은 공산지 중앙부(WS-1) 4,839 cells/mL, 공산지 취수탑(WS-2) 2,269 cells/mL, 덕동호 댐(WS-3) 112 cells/mL, 안계호 취수탑(WS-4) 382 cells/mL, 영천호 취수탑(WS-5) 397 cells/mL, 운문호 댐(WS-6) 70 cells/mL, 운문호 취수탑(WS-7) 70 cells/mL, 진전지 상류(WS-8) 1 cells/mL, 진전지 하류(WS-9) 2 cells/mL로 나타났다. 상수원 지점의 유해남조류 발생도 낙동강 지점과 유사하게 계절적으로 볼 때 5~10월에 발생되고 12~2월 겨울철에는 대부분 지점에서 발생하지 않았다.
3.2. 남조류 종 비율 및 우점종
유해남조류 4종(Microcystis, Anabaena, Oscillatoria, Aphanizomenon)의 우점 비율을 Fig. 4에 나타내었다. 2012~2019년 사이 낙동강 지점(ND-1~ND-8)에서 가장 우점비율이 높았던 종은 Microcystis이었고, 조류 종의 우점 비율은 지점에 따라 Microcystis 47~63%, Anabaena 0~11%, Oscillatoria 0~5%, Aphanizomenon 29~47% 사이로 나타났다(Fig. 4(A)). 낙동강 지점(ND-1~ND-8)은 남조류 종의 비율이 지점별로 큰 차이를 나타내지 않았으나, 하류로 갈수록 Microcystis의 비율이 다소 증가하는 추세를 나타냈다. 낙동강 지점은 우점종의 종류자료만 있고 종별 세포수 자료는 없어서 이에 대한 분석은 할 수 없었다.
상수원 지점(WS-1~WS-9) 중 진전지 상류(WS-8) 지점은 우점종 자료가 없어 제외하고 나머지 지점을 대상으로 분석하였다(Fig. 4(B)). 2012~2019년 사이 상수원 지점(WS-1~WS-9)에서 가장 우점비율이 높았던 종은 지점에 따라 다르게 나타났는데 공산지 중앙부(WS-1), 공산지 취수탑(WS-2), 덕동호 댐(WS-3), 운문호 댐(WS-6), 운문호 취수탑(WS-7), 진전지 하류(WS-9)에서는 Microcystis의 비율이 높았으나, 안계호 취수탑(WS-4)과 영천호 취수탑(WS-5)에서는 Aphanizomenon의 비율이 높았다. 조류 종의 우점 비율은 지점에 따라 Microcystis 14~100%, Anabaena 0~2% Oscillatoria 0~20%, Aphanizomenon 0~76% 사이로 나타났다.
상수원 지점(WS-1~WS-9) 중 안계호와 영천호를 제외하면 Microcystis의 우점 비율은 49% 이상이었다. 종별 세포수가 표기되는 상수원 지점의 자료를 살펴보면 유해남조류 발생량은 대부분이 Microcystis로 나타나 발생 개체수로도 Microcystis가 가장 많았는데, 이는 28~32℃에서 최대 성장률을 보이는 Microcystis가 여름철에 대량 발생하기 때문이다[9]. Microcystis 다음으로 많은 우점 비율을 보인 Aphanizomenon은 23~29℃ 정도의 수온을 선호하는 종이나 타 남조류에 비해 높은 저온 내성을 가지고 있어 늦가을에서 봄철에 주로 우점하였다[10]. 낙동강 지점과 상수원 지점의 우점 횟수를 살펴보면 하천인 낙동강의 경우 대체로 일정한 우점 비율을 보인다는 특징이 있었고 호소인 상수원 지점의 경우 일관성을 보이지 않으며 지점마다 다양한 우점 비율을 나타냈다. 호소의 유해 남조류 발생은 호소별로 개별적인 특성을 보였고 하천과 비교하여 종다양성이 높게 나타났다. 이러한 경향은 금강유역을 대상으로 한 타 연구와도 유사한 결과이다[11].
3.3. 유해남조류 발생과 수질인자의 상관관계
낙동강 지점(ND-1~ND-8)과 상수원 지점(WS-1~WS-9)의 유해남조류 세포수와 수질인자(수온, pH, DO, 클로로필-a) 간의 상관계수를 분석하여 Table 1과 Table 2에 정리하였다. 유해남조류 세포수는 범위가 넓어 log-scale에 대한 상관계수도 함께 분석하였다. 진전지 상류(WS-8) 지점은 2013~2014년 사이 pH와 DO 값이 측정되지 않았고 2015~2019년 사이에는 유해남조류 발생이 없었기에 pH와 DO 항목에 대한 상관계수를 구하지 못하였다.
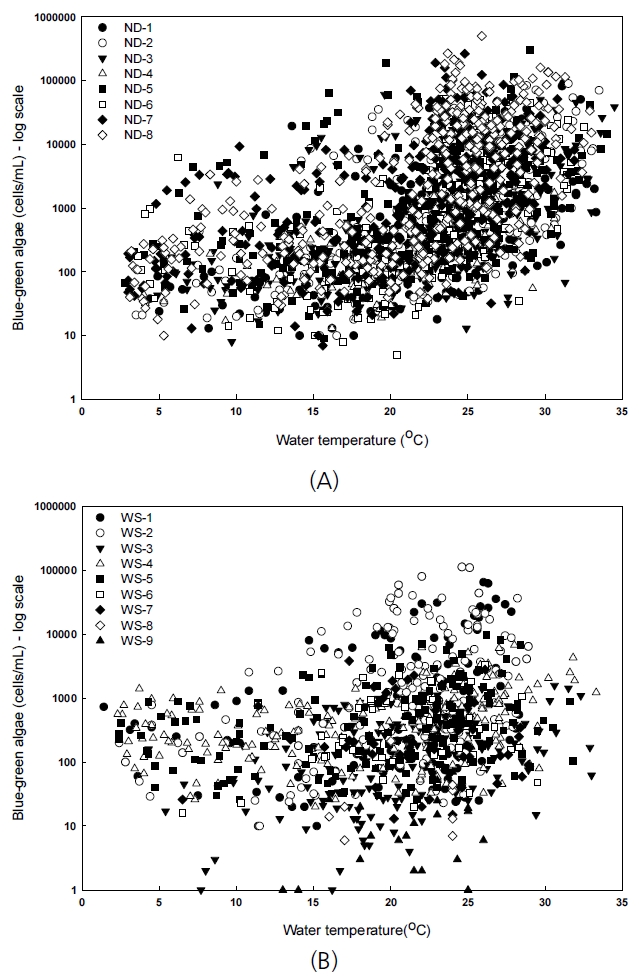
낙동강 지점(ND-1~ND-8) 자료의 경우 유의성이 있는 상관관계를 보이는 것은 log-scale 남조류 세포수와 수온(all data, r=0.71), log-scale 남조류 세포수와 DO(all data, r=-0.66)로 나타났다(Table 1). 낙동강 지점별 남조류 세포수를 log-scale로 표현하면 수온과 0.64~0.77의 양의 상관관계를 보였고, DO 농도와 –0.74~–0.56의 음의 상관관계를 보였다. 즉 수온이 높을 때 남조류가 많이 발생했고, 남조류 생성이 증가할 때 DO 농도는 감소했음을 알 수 있다. 상수원 지점(WS-1~WS-9)에서는 남조류 세포수를 log-scale로 비교해도 수온(r=0.15~0.64) 및 DO(r=–0.52~–0.19)와 낙동강 지점에 비해 낮은 상관관계를 보였다(Table 2). 공산지 지점(WS-1, WS-2)에서만 남조류 세포수와 수온이 상관계수 0.6 이상을 나타냈다.
Fig. 5는 낙동강 지점(ND-1~ND-8)과 상수원 지점(WS-1~WS-9)의 지점별 2015~2019년 유해남조류 발생과 수온의 관계를 나타낸다. 낙동강 지점의 유해남조류 발생은 수온 20℃ 이하에서 대부분 10,000 cells/mL(조류경보 경계단계 기준)를 넘지 않았으나 20℃에서부터 수온이 증가함에 따라 유해남조류 발생이 급격히 증가하는 경향성을 보인다. 상수원 지점에서도 비슷한 경향을 보여 유해남조류 발생이 수온 20℃ 이하에서 대부분 1,000 cells/mL(조류경보 관심단계 기준)를 넘지 않았다. 정도의 차이는 있으나 모든 지점에서 수온이 20℃ 이상으로 증가함에 따라 유해남조류 발생량이 많아지는 경향이 있으므로 수온이 높은 초여름~가을 사이에는 분석 횟수를 늘려 유해남조류 발생 관리 및 예방에 더욱 신경 써야 할 것으로 판단된다.
Fig. 6은 낙동강 지점(ND-1~ND-8)과 상수원 지점(WS-1~WS-9)의 지점별 유해남조류 발생과 DO의 관계를 나타낸다. 상수원 구간에 비해 비교적 유해남조류 발생량이 많은 낙동강 지점에서 유해남조류 발생이 많을수록 DO 농도가 낮아지는 경향을 보였다. 이는 조류발생으로 인해 조류의 광합성을 통한 일시적 DO 농도 상승이 일어날 수 있으나 장기적으로 보면 광합성으로 생성되는 산소보다 호흡으로 사용하는 산소가 더 많고, 수표면을 덮은 조류로 인하여 DO 농도가 낮아짐에도 공기 중의 산소가 물속으로 용해되지 못하는 등 복합적인 이유가 존재하기 때문일 것이다[12]. 또한 남조류 발생 가능성이 높은 여름철에 수온 증가로 용존산소의 포화도가 낮은 것도 이유가 될 수 있다.
그 외 항목인 pH 및 클로로필-a와 남조류 세포수는 낙동강 지점과 상수원 지점 모두 뚜렷한 상관관계가 없는 것으로 확인되었다. 클로로필-a는 남조류 뿐만 아니라 규조류, 녹조류의 개체에도 포함이 되어 있고, 오히려 남조류의 클로로필-a 함유량은 규조류나 녹조류에 비해 낮기 때문으로 판단된다[13,14]. 이에 환경부도 녹조현상과 상관관계가 낮은 클로로필-a 농도를 2016년부터 조류경보제 수질항목에서 삭제하고 남조류 세포수로 단일화하여 관리하고 있다. pH는 유해남조류 개체 성장에 영향을 주는 인자이나 pH 7.7~9.4 범위인 약알칼리성 환경에서만 생장이 잘 일어나고 이 범위를 벗어나면 사멸하거나 활성이 떨어지는 특성을 보이기 때문에 양 혹은 음의 상관계수로는 높은 상관성을 보이지 못하는 것으로 판단된다[15].
한편 본 연구에서는 자료가 미흡하여 분석하지 못했으나 물속의 영양염류, 특히 인(P)의 농도도 조류 세포수와 밀접한 관계가 있다. 미국 환경보호청(USEPA)에 따르면 조류 번성을 억제하기 위한 인의 한계농도는 0.02 mg/L로 알려져 있고[16] 인 농도에 따라 조류 발생이 제어된다는 연구결과도 다수 존재한다[17]. 남조류의 발생은 그 외에도 다양한 수질인자와 유역의 특성에 따라 영향을 받을 수 있고 복잡한 양상을 보임으로[18-24] 단순히 몇 가지 인자로만 설명하기에는 부족하다. 다만, 본 연구의 결과로만 보면 대구・경북지역 낙동강 유역의 남조류 발생은 수온의 영향을 크게 받는 것으로 판단된다.
4. 결 론
조류경보제와 수질예보제로 관리 되고 있는 대구・경북지역 낙동강 수계 및 상수원 지점의 2012년부터 2019년 사이 유해남조류 발생 현황에 대하여 분석한 결과, 아래와 같은 결론을 얻었다.
1) 대구・경북지역 내 조류경보제 및 수질예보제로 관리되고 있는 수계에서 남조류 세포수는 상수원 지점(9개소)보다 낙동강 지점(8개소)에서 더 높게 나타났다.
2) 낙동강 지점의 유해남조류 발생은 하류로 갈수록 정도가 심해지고 보와 보 사이의 지점보다 보 인근에서 더 높은 발생 정도를 보였다.
3) 낙동강 지점에서는 Microcystis가 우점종으로 나타났고 하류로 갈수록 Microcystis의 비율이 다소 증가하는 추세를 나타냈다. 상수원 지점은 지점에 따라 우점비율이 다르게 나타나 Microcystis 또는 Aphanizomenon이 우점종이었다.
4) 유해남조류의 발생은 수온 증가 및 DO 농도 감소와 밀접한 연관이 있었으나, Chl-a는 유해남조류 발생을 예측하는 인자로서 상관성이 낮았다.